
蚊类生物在人类社会中具有重要公共卫生意义,其不仅干扰人畜正常生活,更是登革热、流行性乙型脑炎(乙脑)、疟疾、丝虫病等多种疾病的传播媒介,严重威胁生命健康[1]。在流行病学层面,不同蚊种具有特定的疾病传播特性:白纹伊蚊(Aedes albopictus)和埃及伊蚊(Ae. aegypti)构成登革热的主要传播链;三带喙库蚊(Culex tritaeniorhynchus)是乙脑的核心传播媒介;疟疾的传播涉及雷氏按蚊(Anopheles lesteri)、中华按蚊(An. sinensis)等种类;而丝虫病的传播则与致倦库蚊(Cx. pipiens quinquefasciatus)和淡色库蚊(Cx. pipens pallens)密切相关[2]。作为重要的病媒生物,蚊虫种群数量的动态变化影响传染病的流行趋势。流行病学调查证实,蚊虫密度的异常升高往往伴随着相关疾病的暴发风险,例如登革热与乙脑的流行强度与媒介蚊虫密度呈显著正相关[3]。蚊虫密度是蚊媒传染病的重要危险因素,同时也是蚊虫监测的重要指标[4]。
近年来,随着气候变化的加剧,蚊媒传播疾病的范围和频率也在增加。研究表明,气候变化能够通过改变蚊虫的生态环境,直接影响蚊虫的密度。如温度是影响蚊虫生命周期中每个阶段的关键因素,适宜的温度不仅可以加速蚊虫卵、幼虫和蛹的发育,还能缩短成蚊的生命周期[5];降水量同样也是影响幼蚊孵化和繁殖的重要因素,降水量增加为蚊虫提供了更多的积水环境,使得幼蚊在这些积水中迅速繁殖[6],过多的降雨也可能冲刷蚊虫的孵化场所,从而减少幼蚊的数量[7]。气象因素对媒介生物种群密度的影响往往不会立即显现,而是伴随一定的滞后效应。为了解这一复杂的时滞关系,分布滞后非线性模型(distributed lag non-linear model,DLNM)常被用于相关数据的分析。DLNM作为一种常用的统计工具,能够有效捕捉暴露与健康结果之间的非线性和滞后效应[8],尤其适用于处理气象因素对蚊虫种群密度的影响。
重庆市地处中国西南部,具有典型的亚热带季风气候特征,夏季高温且降水丰富,这为蚊虫的繁殖和活动提供了良好的气候条件。然而,过往针对重庆地区气候因素滞后效应对蚊虫密度影响的研究有限,因此本研究采用DLNM模型分析重庆市2013-2022年4-11月气象数据对相应月份幼蚊密度的影响,以确定影响幼蚊密度的关键气象指标和滞后效应,为重庆市预测蚊虫密度变化和防控登革热等重要蚊媒传染病提供科学的参考依据。
1 材料与方法
1.1 蚊密度数据
本研究的幼蚊密度数据由重庆市疾病预防控制中心提供。根据《全国病媒生物监测实施方案》(中疾控传防发〔2016〕56号),采用路径法,每年4-11月开展监测,每月中旬进行1次,全年共监测8次,覆盖重庆市35个县(区),每个县(区)作为1个监测点。因每年12月到次年3月未对蚊密度进行监测,故对应月份的幼蚊密度数据和气象数据未被纳入研究。各监测点以人居环境为核心,覆盖居民区(500 m)、有独立院落单位(500 m)、公园(300 m)、废旧轮胎集中堆放地(200 m)、道路/沿街绿带(500 m)(均指延长米)等5类场所外环境,记录路径沿线小型积水容器总数及其中阳性容器积水(检出伊蚊幼蚊或蛹)数量,用以计算路径指数。
路径法通过计算路径指数(path index,PI)反映幼蚊密度状况[9]。
PI(处/km)=(阳性容器数+阳性小型积水数)(处)/监测行走距离(km)。
1.2 气象数据
本研究所用气象数据来自美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration,NOAA)1942-2024年中国观测站点逐月气象指标数据集。下载重庆市沙坪坝气象站(站点编号:57516)2013-2022年每年4-11月的气象信息,包括月平均降水量(PRCP)、平均气温(TEMP)、露点(DEWP)、海平面气压(SLP)、站点气压(STP)、平均风速(WDSP)、最大风速(MXSPD)、最高气温(MAX)、最低气温(MIN)9个指标。
1.3 统计分析方法
对每月幼蚊密度和气象数据进行描述性统计,利用R 4.4.0中基于局部加权回归(locally weighted scatterplot smoothing,LOESS)的季节趋势分解模型(seasonal and trend decomposition using LOESS,STL)对逐月幼蚊密度时间序列数据进行差分,分析其季节性趋势和长期趋势;对幼蚊密度与各气象因素进行Spearman秩相关分析,探讨各气象因素与幼蚊密度的相关性,确定可以纳入模型的气象因素。为避免共线性问题,当两气象因素相关性过高(ρ > 0.8)时不同时纳入模型。本研究使用“dlnm”包构建DLNM模型,同时利用“splines”包实现自然样条函数。所使用的DLNM模型的建模核心思想是交叉基,利用交叉基函数分析气象因素与幼蚊密度之间的非线性关系及滞后效应,对变量建立交叉基函数并引入广义线性模型(general linear model,GLM),最终确定其模型结构为:
μt为每月幼蚊密度均值,g()为连接函数,由于捕获幼蚊相对种群总数为小概率事件,故选择类泊松模型为连接函数。a为截距,cb()为气象因子的交叉基函数,weather为最终纳入模型的气象因子,lag为最长时间;ns()为自然样条函数,time为时间变量,df为对应的自由度,ns(time, df)用来控制长期趋势和季节性效应;β和month为月份变量及其回归系数,用于捕捉未被时间变量完全解释的月份特异性效应。采用GLM模型和自然样条函数探究降水量、气温与幼蚊密度之间的非线性关系,确定DLNM的各个参数。改变纳入模型气候变量的自由度(2~5)、最大滞后时长(1~2月)及滞后时长自由度(2~3)、时间变量time自由度(5~10),将不同参数组合多次构建模型并计算各个模型的广义赤池信息准则(quasi-Akaike information criterion,QAIC),以QAIC最小值确定最优模型。经过反复验证,最终确定以QAIC值为80.57的模型为最终模型,分析气象因素滞后效应对幼蚊密度的影响。确定月平均气温、月平均降水量的df=3,滞后变量的自由度参数设置为3,月平均气温和月累计降水量的时间变量自由度均设置为8,最长滞后时长设置为2个月。
2 结果
2.1 幼蚊密度基本特征
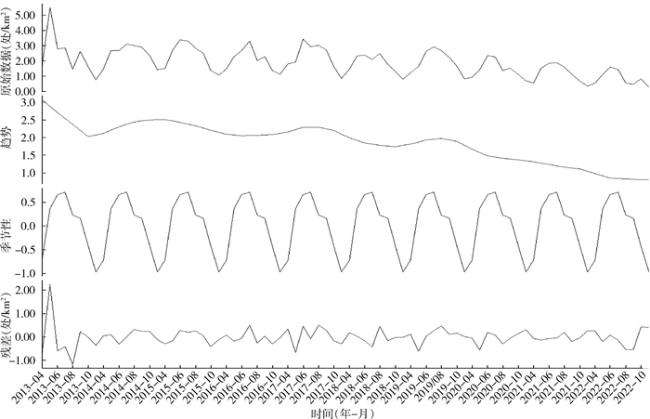
2013-2022年重庆市幼蚊密度每月的平均PI值为1.86,PI值的中位数为1.62。对研究期间的幼蚊密度时间序列进行STL模型分解,见图 1。从长期趋势来看,2013-2022年幼蚊密度指数总体呈现明显的下降趋势。幼蚊密度变化呈现明显的季节性变化特征,每年4-6月季节指数逐渐升高,在6月达到峰值后逐渐下降。
2.2 气象因素基本特征
研究期间月平均降水量、月平均气温、月平均露点、月平均海平面气压、月平均站点气压、月平均风速、月平均最大风速、月平均最高气温、月平均最低气温的中位数分别为142.84 mm、22.97 ℃、18.38 ℃、1 009.27 hPa、979.81 hPa、4.26 m/s、8.85 m/s、26.91 ℃、19.66 ℃,其他基本特征见表 1。
表1 重庆市2013-2022年4-11月各气象因素的基本特征Tab. 1 Basic characteristics of meteorological factors in Chongqing, China, April to November, 2013-2022 |
| 气象因素 | 均值 | 标准差 | 最小值 | 最大值 | P5 | P25 | P50 | P75 | P95 |
| 累计降水量(mm) | 149.56 | 69.81 | 27.18 | 339.42 | 43.78 | 97.15 | 142.84 | 189.57 | 276.77 |
| 气温(℃) | 22.97 | 5.26 | 11.99 | 34.35 | 13.60 | 19.33 | 22.96 | 27.14 | 31.02 |
| 露点(℃) | 17.57 | 7.18 | 9.45 | 23.78 | 10.26 | 14.82 | 18.38 | 20.95 | 22.87 |
| 海平面气压(hPa) | 1 009.43 | 6.70 | 998.60 | 1 023.26 | 1 000.05 | 1 002.82 | 1 009.26 | 1 015.14 | 1 021.44 |
| 站点气压(hPa) | 981.33 | 10.06 | 959.69 | 1 021.52 | 971.58 | 974.11 | 979.81 | 986.68 | 1 002.12 |
| 风速(m/s) | 4.25 | 0.44 | 3.25 | 5.67 | 3.50 | 3.98 | 4.26 | 4.49 | 4.98 |
| 最大风速(m/s) | 8.90 | 0.92 | 6.90 | 11.14 | 7.29 | 8.27 | 8.84 | 9.64 | 10.46 |
| 最高温度(℃) | 27.42 | 10.75 | 15.56 | 40.91 | 16.13 | 23.53 | 26.91 | 32.31 | 23.36 |
| 最低温度(℃) | 19.33 | 8.56 | 8.76 | 29.08 | 11.46 | 15.89 | 119.66 | 36.08 | 26.56 |
2.3 蚊虫密度与气象因素的相关性分析
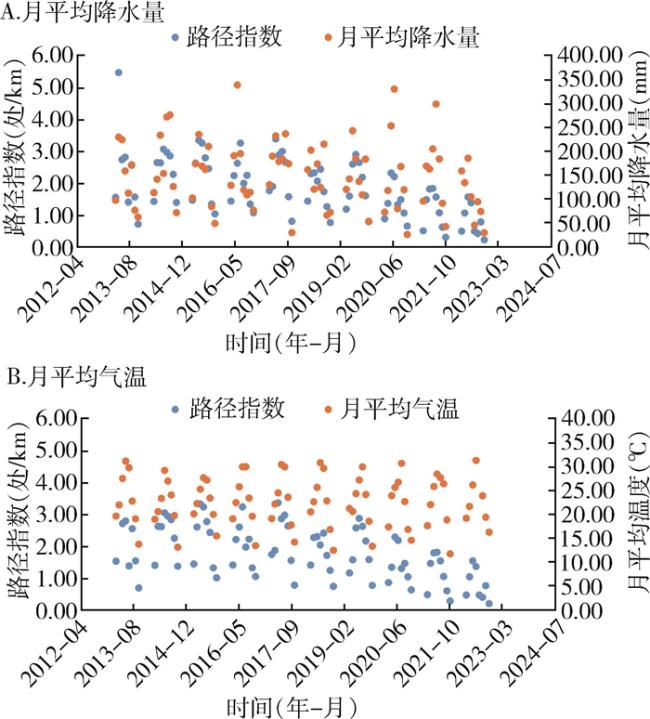
蚊虫的PI与月平均降水量、月平均温度、月平均露点、月平均最高温度、月平均最低温度呈正相关,相关系数分别为0.75、0.59、0.63、0.51、0.62;与月平均海平面气压、月平均站点气压呈负相关,相关系数分别为-0.62和-0.47;与月平均风速、月平均最大风速之间的相关关系无显著性(均P > 0.05),见图 2。为排除不同气象因素之间的共线性,使用方差膨胀因子(variance inflation factor,VIF)来评估共线性问题,VIF值越高,表明共线性越强,通常VIF > 10.000表示共线性较强,最终选择月平均降水量和月平均温度作为纳入模型的气象因素,其VIF值分别为1.349和1.336。月平均气温和降水量与幼蚊密度在时间分布上同步波动,数据具时序性特征,可用于后续构建模型,见图 3A、3B。
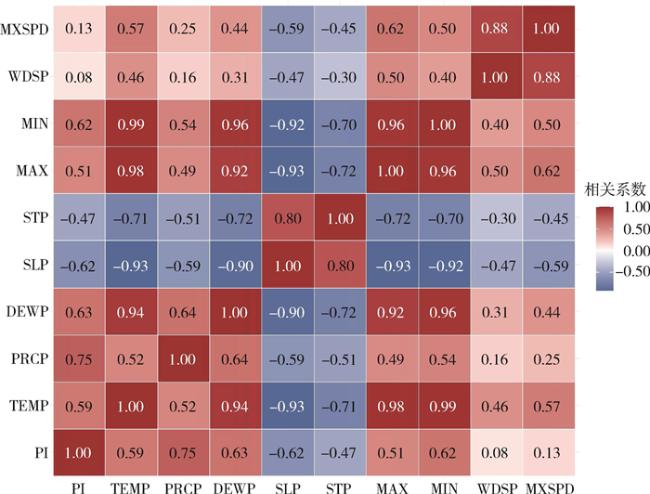
图2 重庆市2013-2022年4-11月气象因素与幼蚊密度相关性注:蓝色表示相关系数为负值,红色表示相关系数为正值;MAX最高气温;MXSPD最大风速;WDSP平均风速;STP站点气压;SLP海平面气压;DEWP露点;TEMP平均气温;PRCP月平均降水量;MIN最低气温;PI幼蚊路径指数。 Fig. 2 Correlation between meteorological factors and mosquito larvae density in Chongqing, China, April to November, 2013-2022 |
2.4 气象因素对幼蚊密度滞后效应分析
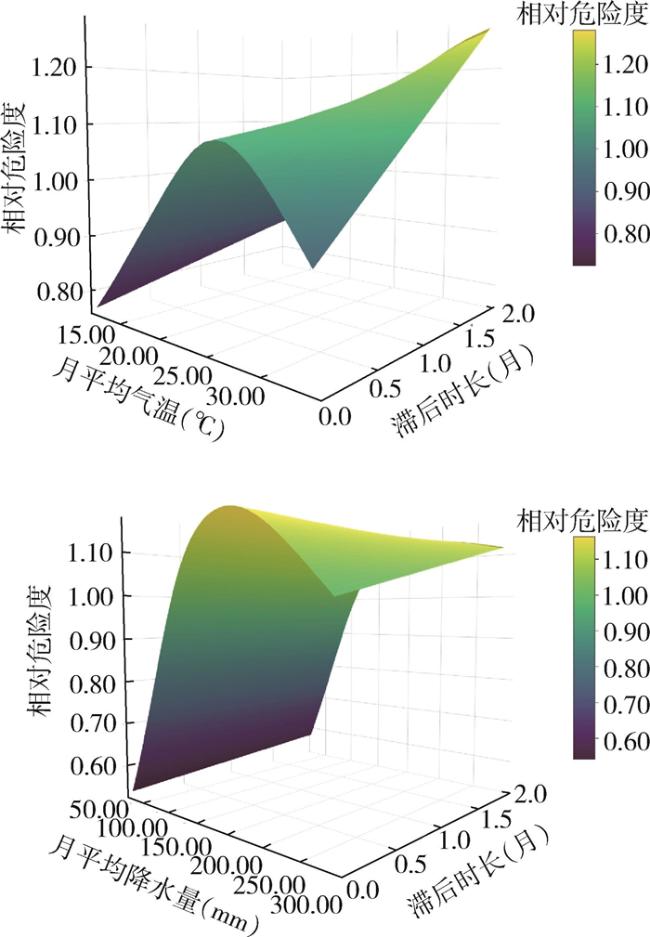
以节点月平均温度20.96 ℃和月平均降水量114.78 mm为参考值,将月累计降水量、月平均气温与滞后时长组合构建三维效应图,分析不同滞后时长下纳入的气象因素对幼蚊密度的效应,见图 4。
2.4.1 月平均气温对幼蚊密度的滞后效应
2013-2022年的4-11月,重庆市月平均气温变化范围为11.99~34.35 ℃,月平均气温对同期幼蚊密度的危险效应不显著(P > 0.05)。滞后0.2月(约6 d)时,相对危险效应在20.00~23.02 ℃显著(P < 0.05),随气温升高而增加,在月平均温度达到23.02 ℃时相对危险度(relative risk,RR)值最高(RR=1.06,95%CI:1.00~1.13)。滞后0.4月(约12 d)时,相对危险效应在16.53~31.38 ℃显著(P < 0.05),相对危险效应随气温升高逐渐增加,在29.98 ℃达到峰值(RR=1.18,95%CI:1.03~1.35),之后随气温的继续升高,相对危险效应逐渐降低,在月平均气温达到31.38 ℃时相对危险效应降至最低点(RR=1.17,95%CI:1.02~1.34)。滞后0.6月(约18 d)时,相对危险效应在29.43~30.66 ℃显著(P < 0.05),RR值随月平均气温的升高而逐渐上升,在月平均气温为30.66 ℃时,RR值最高(RR=1.23,95%CI:1.00~1.51)。月平均气温变化对幼蚊密度的滞后危险效应在滞后时长0.3月(约18 d)之后不显著(P > 0.05)。见图 5。
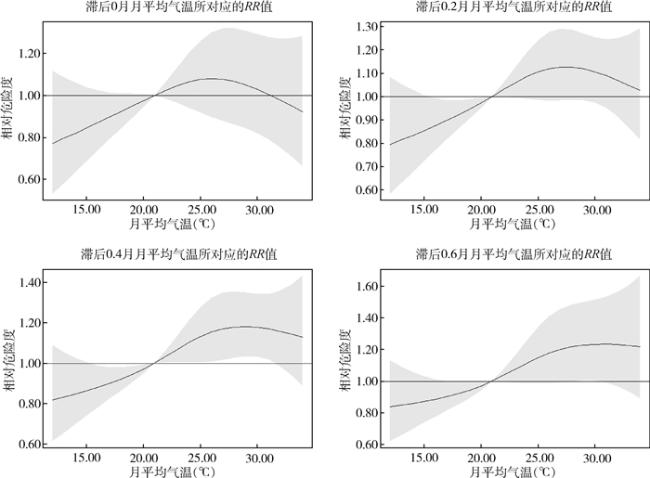
2.4.2 月平均降水量对幼蚊密度的滞后效应
2013-2022年重庆市4-11月的月平均降水量变化范围为27.18~339.42 mm,月平均降水量在27.18~243.91 mm时,同期的幼蚊密度受到显著影响(P < 0.05),随着月平均降水量的增加,RR值先上升后下降,最大值出现在月平均降水量为203.23 mm时(RR=1.16,95%CI:1.04~1.30),之后,随着降水量的进一步增加,RR值逐渐递减。滞后0.2月(约6 d)时,降水量在27.18~276.99 mm时对幼蚊密度的影响显著(P < 0.05),RR值在210.70 mm时达到最大值(RR=1.16,95%CI:1.05~1.29)。滞后0.4月(约12 d)时,相对危险效应在27.18~276.99 mm时显著(P < 0.05),最大RR值出现在215.91 mm时(RR=1.16,95%CI:1.04~1.29)。滞后0.6月(约18 d)时,显著影响出现在27.18~285.00 mm(P < 0.05),RR峰值在225.03 mm处(RR=1.16,95%CI:1.03~1.30)。滞后0.8月(约24 d)时,降水量在27.18~272.69 mm时相对危险效应显著(P < 0.05),RR值在243.91 mm时达到峰值(RR=1.15,95%CI:1.01~1.31)。滞后1个月(约30 d)时,相对危险效应在27.18~203.23 mm时显著(P < 0.05),RR值在203.23 mm时达到峰值(RR=1.13,95%CI:1.00~1.28)。滞后1.2月(约36 d)时,相对危险效应在27.18~175.11 mm时显著(P < 0.05),RR值在175.11 mm时达到峰值(RR=1.10,95%CI:1.00~1.21)。滞后1.4月(约42 d)时,相对危险效应在27.18~168.97 mm时显著(P < 0.05),最大RR值出现在168.97 mm时(RR=1.08,95%CI:1.00~1.17)。在滞后1.4月(约42 d)之后,月平均降水量对幼蚊密度的滞后影响不再显著(P > 0.05)。见图 6。
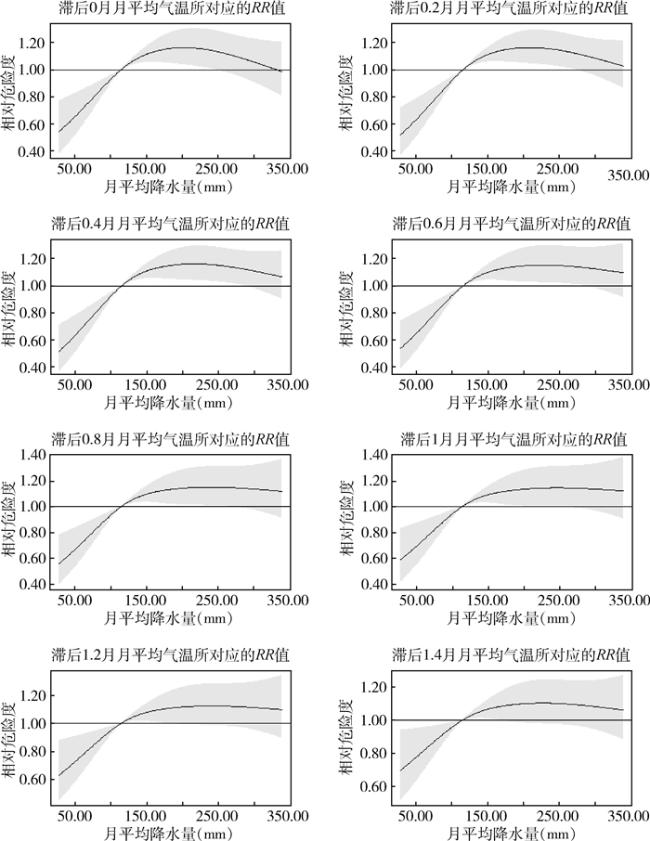
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.5 敏感性分析
为验证DLNM模型参数设置对结果的影响,在主要模型结果的基础上,分别将月平均气温和月平均降水量2个变量的自由度调整为2~5,同时考察时间变量自由度在6~10的影响。结果显示,不论参数如何调整,温度与降水量对幼蚊密度的滞后效应趋势“暴露-滞后-反应”的关联变化不大,表明模型结果较稳健。
3 讨论
气候因素,特别是温度和降水量,对幼蚊密度具有显著影响。它们通过影响蚊虫的繁殖、孵化速度以及幼蚊的发育过程,进而改变幼蚊的密度。此外,这些气象因素对幼蚊的发育和密度变化并非立竿见影,而是需要一定周期才能发挥作用,可能经过数周甚至数月后,才会在蚊密度的变化中体现出来,因此导致了滞后效应的出现[10]。本研究分析了选用的气象因素与幼蚊密度的关联,并利用DLNM模型,探讨最终纳入的月平均降水量和月平均气温2个变量与幼蚊密度之间存在的非线性关系以及滞后效应。
研究发现,重庆市月平均气温变化对幼蚊密度不会立即产生影响,而是存在滞后效应,最长滞后效应时间为0.6月(约18 d),在关于气候与蚊密度的研究中,温度变化对幼蚊密度的滞后效应通常在2~3周(14~21 d)显现出来[11],这与18 d的最长滞后期相符合。月平均温度范围在27~31 ℃,幼蚊密度较高,这个温度范围与幼蚊的最适生长温度(28~30 ℃)相符合[12],在30.66 ℃、滞后0.6月(约18 d)时RR达到峰值,随着温度进一步升高,RR值呈现下降的趋势。既往研究表明,温度是影响幼蚊发育速度、存活率以及繁殖成功的关键因素,在幼蚊的最适发育温度范围内,幼蚊从孵化到成蚊的发育时间显著缩短,存活率较高,但是当温度 > 35 ℃时,发育会受到抑制,甚至导致较高的死亡率[13],从而导致温度对幼蚊密度的RR值逐渐降低,甚至 < 1.00。
重庆市月平均降水量对幼蚊密度存在显著的滞后效应,最长滞后效应时间为1.4月(约42 d),这与气象因素对埃及伊蚊幼蚊密度影响的研究结果基本吻合[14]。本研究发现,滞后时长在0~0.8月内(约0~24 d),在月平均降水量可对幼蚊密度产生显著影响的范围内,RR值随着降水量的增加先上升,达到峰值后逐渐下降,与滞后时间呈现出倒“U”型关系的趋势;滞后时长0.8~1.4月(约24~42 d),RR值随降水量的增加而增加;滞后0.8月(约24 d)、月平均降水量243.91 mm时出现最大相对危险效应。在短滞后期,降水可以为幼蚊提供适合的孳生地,特别是中小型积水容器如瓶罐、沟渠等,容积适中的水体有利于伊蚊卵的孵化以及幼蚊的生长[15]。随着降水的持续增加,积水逐渐饱和,适合的孳生地达到最大容量,RR值随之达到峰值;当降水量过多时,强降雨可能会导致积水被冲刷、干扰和破坏[16],或者因水体过深影响水温和氧气含量[17],进而抑制蚊卵孵化,导致RR值下降。在较长滞后期,RR值随降水量增加而上升,可能是由于一些持久性孳生地长时间维持了适合的积水量,此时降水的累积效应更为明显[18],这些水体不仅能维持蚊虫发育周期,还能够在更长的滞后时段中继续提供孳生环境[19],促进幼蚊数量的累计增长。
从防控角度来看,本研究结果对重庆市乃至更广泛地区的蚊媒密度监测和预警具有重要意义。结合气象预测数据,可在幼蚊密度预计升高前采取针对性防控措施:在气温接近30 ℃或降水量 > 200 mm的滞后时段内,应加强重点区域的环境治理、开展定点监测和精准投药,以降低蚊媒密度,从而减少登革热等蚊媒传染病的暴发风险。此外,长期监测应结合动态调整的防控策略,以适应气候变化对蚊媒生态的潜在影响。
本研究也存在一定的局限性:本研究基于重庆市全市范围内的数据进行分析,未能考虑到不同区域间的空间异质性,这可能影响到对局部地区蚊密度变化的精准预测;研究未考虑干预活动如健康宣讲、病媒生物控制对幼蚊密度的影响;研究主要关注单独的气象因素对幼蚊密度的影响,而未能综合考虑气象因素之间及气象因素与非气象因素(如社会经济条件、城市化进程、病媒控制措施等)间的交互作用,在实际情况中可能是多个因素同时和交互对蚊虫生长发育产生影响。
综上所述,本研究采用DLNM模型探讨了气象因素,特别是月平均气温和月平均降水量对重庆市2013-2022年幼蚊密度的滞后效应,这一滞后效应的分析为理解气象因素变化对蚊密度的影响提供了科学依据。未来应进一步深化对气象-蚊媒-疾病传播的系统研究,以优化防控策略,提高公共卫生应对能力。