
登革热(dengue fever,DF)是一种由登革病毒(Dengue virus,DENV)引起的急性蚊媒传染病,主要传播媒介是埃及伊蚊(Aedes aegypti)和白纹伊蚊(Ae. albopictus)。自2013年以来登革热在云南省德宏傣族景颇族自治州(德宏州)、西双版纳傣族自治州(西双版纳州)和临沧市等边境地区多次暴发,已成为这些地区重要的蚊媒传染病,严重危害人民群众健康[1]。景洪市隶属云南省西双版纳州,属热带和南亚热带湿润季风气候区,适合蚊虫孳生,埃及伊蚊和白纹伊蚊同域分布,且埃及伊蚊是优势伊蚊种群[2]。2013-2023年景洪市共暴发了5次大规模登革热疫情,特别是2023年暴发了历史上最大的1次疫情,报告病例3 381例[3]。因此,登革热是景洪市重要的公共卫生问题,引起严重经济负担和健康危害。快速有效降低埃及伊蚊或白纹伊蚊密度是登革热防制的重要手段。除了卫生杀虫剂的使用,利用沃尔巴克氏体(Wolbachia)控制蚊虫的繁殖和预防疾病的传播给蚊媒传染病的预防控制工作带来了新的思路。昆虫共生菌沃尔巴克氏体隶属变形菌门(Proteobacteria)α变形菌亚纲(Alphaproteobacteria)立克次体目(Rickettsiales)立克次体科(Rickettsiaceae)的沃尔巴克氏体属,是一类通过母系遗传且天然存在却无法感染人或其他脊椎动物的革兰阴性细胞内共生细菌,广泛存在于节肢动物尤其是昆虫纲动物中[4-5]。沃尔巴克氏体通过胞质不相容(cytoplasmic incompatibility,CI)、孤雌生殖(parthenogenesis)、雌性化(feminization)等多种方式调控昆虫生殖[6]。依据沃尔巴克氏体的CI特性,人们制定出种群压制和种群替换策略,并研究出了沃尔巴克氏体诱导的昆虫不相容技术(incompatible insect technique,IIT)和病原阻断技术(pathogen blocking technique,PBT)用于蚊媒传染病的防控[7]。奚志勇等[8]2014年采用白纹伊蚊沃尔巴克氏体wPip、wAlbA和wAlbB株系三重感染品系,成功抑制野外蚊虫种群增长,野外蚊虫种群卵孵化率和成虫数量降低达94%。研究表明蚊虫可自然携带沃尔巴克氏体,对沃尔巴克氏体运用于蚊媒传染病防控产生影响[9]。本研究对2023年云南省景洪市埃及伊蚊和白纹伊蚊种群自然感染沃尔巴克氏体进行调查研究,旨在为运用沃尔巴克氏体控制蚊媒传染病提供科学依据。
1 材料与方法
1.1 样本来源
2023年在云南省景洪市东、南、西、北、中5个不同方位,采集伊蚊幼虫,同一孳生容器内伊蚊幼虫收集于同一个采集瓶内,带回实验室,随机吸取幼虫,在100倍显微镜下依据形态学特征进行种类鉴定[10]。鉴定为埃及伊蚊孳生的容器,每个容器内随机抽取≥8只埃及伊蚊幼虫,放置于含有75%乙醇溶液的冻存管中,置于-80 ℃保存。每个容器内剩余伊蚊幼虫,在实验室条件下饲养至羽化后,收集埃及伊蚊雌蚊、雄蚊各≥8只,保存于75%乙醇溶液中,并置于4 ℃保存。由于白纹伊蚊幼虫多为室外容器孳生,采集难度较大,难以在5个方位的同一孳生容器内收集足够多的幼虫,故白纹伊蚊样本采集为景洪市5个方位随机抽取,共获取白纹伊蚊幼、雌和雄蚊各32只,保存方法同埃及伊蚊。样本信息见表 1。
表1 云南省景洪市野生种群伊蚊采集信息Tab. 1 Information on wild Aedes populations in Jinghong, Yunnan Province |
| 蚊种 | 方向 | 采集地点 | 雌蚊(只) | 雄蚊(只) | 幼蚊(只) | 地理坐标 | 采集时间 |
| 埃及伊蚊 | 东 | 告庄景兰寨 | 16 | 16 | 16 | 22°01'14″N,100°81'68″E | 2023年9月 |
| 告庄景罕寨 | 16 | 16 | 16 | 22°01'15″N,100°81'73″E | 2023年9月 | ||
| 南 | 曼龙匡 | 16 | 16 | 16 | 21°99'06″N,100°81'07″E | 2023年9月 | |
| 曼弄枫 | 16 | 16 | 16 | 21°97'22″N,100°79'85″E | 2023年9月 | ||
| 西 | 万达三期 | 16 | 16 | 16 | 22°02'16″N,100°75'81″E | 2023年9月 | |
| 花卉园小区 | 16 | 16 | 16 | 22°00'96″N,100°78'29″E | 2023年9月 | ||
| 北 | 曼斗村 | 16 | 16 | 16 | 22°01'55″N,100°81'02″E | 2023年8月 | |
| 鑫顺鑫北城 | 16 | 16 | 16 | 22°01'80″N,100°81'20″E | 2023年10月 | ||
| 中 | 景洪城区 | 16 | 16 | 16 | 22°01'65″N,100°79'31″E | 2023年10月 | |
| 龙舟广场 | 16 | 16 | 16 | 22°00'31″N,100°81'06″E | 2023年10月 | ||
| 白纹伊蚊 | - | 景洪 | 32 | 32 | 32 | - | 2023年10月 |
注:- 白纹伊蚊为多采样点混合样品,未记录经纬度信息。 |
1.2 蚊虫基因组DNA提取及wsp基因扩增和测序
收集的伊蚊样本按照核酸提取试剂盒[天根生化科技(北京)有限公司,批号:23040610T324]说明书,单只蚊提取基因组DNA。按照文献[11-12]合成wsp基因通用引物(81F-GGTCCAATAAGTGATGAAGAAAC,691R-AAAAATTAAACGCTACTCCA,扩增产物长度为590~632 bp)。PCR反应体系:包含2×Taq预混液12.5 μl,10 μmol/L正、反引物各1.0 μl,DNA模板2.0 μl,加双蒸灭菌水至25.0 μl。wsp通用引物扩增反应条件为:94 ℃ 3 min;94 ℃ 30 s,55 ℃ 1 min,72 ℃ 1 min,35个循环;72 ℃后延伸7 min。Wolbachia的wsp扩增阳性样本,继续进行wAlbA和wAlbB亚群基因(wAlbA和wAlbB)片段扩增(wAlbA扩增引物为328F-CCAGCAGATACTATTGCG,691R-AAAAATTAAACGCTACTCCA,产物长度约350 bp,53 ℃退火45 s;wAlbB扩增引物为183F-AAGGAACCGAAGTTCATG,691R-AAAAATTAAAC GCTACTCCA,产物长度约480 bp,72 ℃延伸31 s。反应体系和其他反应条件同wsp扩增)。扩增产物经1.5%琼脂糖凝胶电泳检测,凝胶成像分析系统观察电泳条带大小,将目的条带大小相符的样品送生工生物工程(上海)股份有限公司昆明分公司测序。
1.3 系统发育分析
用SnapGene 6.0.2软件观察所测序列的峰图形状和测序质量,判断结果的可靠性,并在美国国家生物技术信息中心(NCBI)数据库使用基于局部比对算法的搜索工具(basic local alignment search tool,BLAST)进行同源性比对,利用MEGA 11软件进行DNA序列的多重序列比对,采用邻接(neighbor-joining,NJ)法构建系统发育树(Bootstrap值为1000)。DnaSP 5软件计算单倍型,Popart 1.7软件构建中间系统发育网络(median-joining network)图,即单倍型网络图。
1.4 统计学分析
使用WPS Office 12.1软件对数据进行录入、整理,计算埃及伊蚊和白纹伊蚊的沃尔巴克氏体自然感染率,采用SPSS 27.0软件进行数据分析,采用χ2检验或Fisher确切概率法比较景洪市不同方位、不同虫态和性别埃及伊蚊和白纹伊蚊的沃尔巴克氏体自然感染率。检验水平α=0.05。
2 结果
2.1 埃及伊蚊和白纹伊蚊沃尔巴克氏体自然感染比较
共检测埃及伊蚊DNA样本480份,其中雌、雄和幼蚊各160份,wsp基因阳性样本共计17份,沃尔巴克氏体自然感染率为3.54%(17/480)。东、南、西、北、中5个方位的埃及伊蚊沃尔巴克氏体自然感染率为0~6.25%,西方位采集的埃及伊蚊未检出沃尔巴克氏体感染,见表 2,Fisher确切概率法比较不同方位沃尔巴克氏体自然感染率差异无统计学意义(P=0.132)。共检测白纹伊蚊样本96份,其中雌、雄和幼蚊各32份,白纹伊蚊阳性样本共计89份,沃尔巴克氏体自然感染率为92.71%,见表 3。χ2检验埃及伊蚊和白纹伊蚊沃尔巴克氏体自然感染率差异有统计学意义(χ2=423.581,P < 0.001),白纹伊蚊沃尔巴克氏体自然感染率高于埃及伊蚊。
表2 云南省景洪市不同方位、虫态和性别埃及伊蚊沃尔巴克氏体自然感染情况Tab. 2 The status of natural infection with Wolbachia in Aedes aegypti by geographical orientation, developmental stage, and sex in Jinghong, Yunnan Province |
| 采集 方位 | 雌蚊 | 雄蚊 | 幼蚊 | 总计 | |||||||
| 检出样本数(份) | 检出率(%) | 检出样本数(份) | 检出率(%) | 检出样本数(份) | 检出率(%) | 检出样本数(份) | 检出率(%) | ||||
| 东 | 1 | 3.13 | 4 | 12.50 | 0 | 0.00 | 5 | 5.21 | |||
| 南 | 1 | 3.13 | 1 | 3.13 | 0 | 0.00 | 2 | 2.08 | |||
| 西 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | |||
| 北 | 1 | 3.13 | 1 | 3.13 | 2 | 6.25 | 4 | 4.17 | |||
| 中 | 1 | 3.13 | 1 | 3.13 | 4 | 12.50 | 6 | 6.25 | |||
| 合计 | 4 | 2.50 | 7 | 4.38 | 6 | 3.75 | 17 | 3.54 | |||
注:不同采集方位雌、雄和幼蚊的检测样本数量均为32只。 |
表3 云南省景洪市不同虫态、性别埃及伊蚊和白纹伊蚊wAlbA,wAlbB及wAlbA+wAlbB自然感染情况Tab. 3 The status of natural infection with wAlbA, wAlbB, and wAlbA+wAlbB in Aedes aegypti and Ae. albopictus by developmental stage and sex in Jinghong, Yunnan Province |
| 蚊种 | 虫态/性别 | 检测数 (份) | 检出率 (%) | wAlbA | wAlbB | wAlbA+wAlbB | |||||
| 检出样本数 (份) | 构成比 (%) | 检出样本数 (份) | 构成比 (%) | 检出样本数 (份) | 构成比 (%) | ||||||
| 埃及伊蚊 | 雌蚊 | 4 | 2.50 | 3 | 75.00 | 0 | 0.00 | 1 | 25.00 | ||
| 雄蚊 | 7 | 4.38 | 0 | 0.00 | 1 | 14.29 | 6 | 85.71 | |||
| 幼蚊 | 6 | 3.75 | 3 | 50.00 | 2 | 33.33 | 1 | 16.67 | |||
| 合计 | 17 | 3.54 | 6 | 35.29 | 3 | 17.65 | 8 | 47.06 | |||
| 白纹伊蚊 | 雌蚊 | 31 | 96.86 | 3 | 9.68 | 1 | 3.22 | 27 | 87.10 | ||
| 雄蚊 | 29 | 90.63 | 3 | 10.34 | 1 | 3.45 | 25 | 86.21 | |||
| 幼蚊 | 29 | 90.63 | 3 | 10.34 | 0 | 0.00 | 26 | 89.66 | |||
| 合计 | 89 | 92.71 | 9 | 10.11 | 2 | 2.25 | 78 | 87.64 | |||
注:检测数即各类型样本中的沃尔巴克氏体阳性数,检出率即沃尔巴克氏体检出率。 |
2.2 不同虫态和性别埃及伊蚊和白纹伊蚊沃尔巴克氏体自然感染
埃及伊蚊成蚊沃尔巴克氏体自然感染率为3.44%(11/320),幼蚊为3.75%(6/160)。χ2检验成蚊和幼蚊沃尔巴克氏体自然感染率差异无统计学意义(χ2=0.030,P=1.000);雌蚊沃尔巴克氏体自然感染率为2.50%(4/160),雄蚊为4.38%(7/160),不同性别埃及伊蚊沃尔巴克氏体自然感染率差异无统计学意义(χ2=0.847,P=0.357)。见表 2。
白纹伊蚊成、幼蚊的沃尔巴克氏体自然感染率分别为93.75%(60/64)和90.63%(29/32),χ2检验二者自然感染率差异无统计学意义(χ2=0.308,P=0.579);雌蚊沃尔巴克氏体自然感染率为96.86%(31/32),雄蚊为90.63%(29/32),不同性别沃尔巴克氏体自然感染率差异无统计学意义(χ2=1.067,P=0.302)。见表 3。
比较埃及伊蚊和白纹伊蚊成蚊沃尔巴克氏体自然感染率,差异有统计学意义(χ2=288.638,P < 0.001),其中埃及伊蚊和白纹伊蚊雌蚊沃尔巴克氏体自然感染率差异有统计学意义(χ2=185.350,P < 0.001),埃及伊蚊和白纹伊蚊雄蚊沃尔巴克氏体自然感染率差异有统计学意义(χ2=185.492,P < 0.001)。比较埃及伊蚊和白纹伊蚊幼蚊沃尔巴克氏体自然感染率,差异有统计学意义(χ2=135.018,P < 0.001)。
2.3 埃及伊蚊和白纹伊蚊wAlbA+wAlbB共感染比较
基于wsp通用引物扩增阳性的106份样本,PCR法进行wAlbA和wAlbB特异性分型,结果显示,埃及伊蚊自然感染沃尔巴克氏体的17份样本中仅wAlbA感染6份,仅wAlbB感染3份,wAlbA+wAlbB共感染8份,共感染样本构成比为47.06%(8/17)。白纹伊蚊自然感染沃尔巴克氏体的89份样本中仅wAlbA感染9份,仅wAlbB感染2份,wAlbA+wAlbB共感染78份,共感染样本构成比为87.64%(78/89)。埃及伊蚊和白纹伊蚊wAlbA+wAlbB共感染构成比差异有统计学意义(χ2=15.356,P < 0.001)。见表 3。
2.4 沃尔巴克氏体wsp基因序列系统发育分析
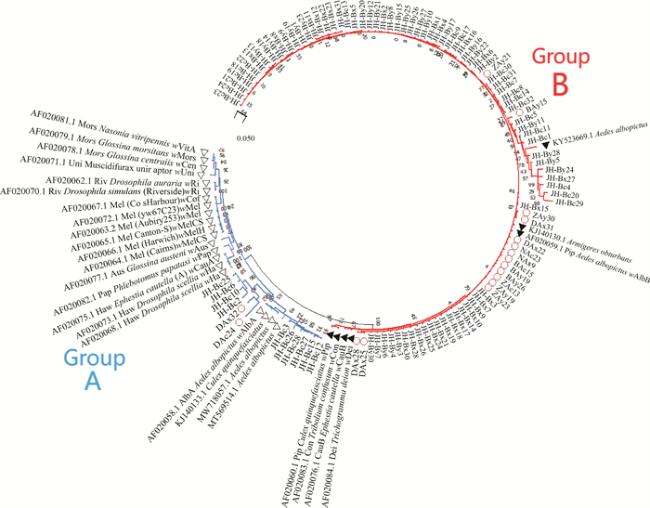
将从埃及伊蚊和白纹伊蚊中获得的106条沃尔巴克氏体wsp基因序列与GenBank数据库中下载的28条已知沃尔巴克氏体基因序列,包含A超群8个亚群(GenBank登录号:AF020081.1、AF020079.1、AF020078.1、AF020071.1、AF020062.1、AF020070.1、AF020067.1、AF020072.1、AF020063.1、AF020065.1、AF020066.1、AF020064.1、AF020077.1、AF020082.1、AF020075.1、AF020073.1、AF020068.1、AF020058.1、KJ140133.1、MW718057.1、MT569514.1),以及B超群4个亚群(GenBank登录号:AF020060.1、AF020083.1、AF020076.1、AF020084.1、AF020059.1、KJ140130.1、KY523669.1)序列,共同进行系统发育分析,采用邻接法构建进化树。所有序列聚成2个大支,分别归入A超群(蓝色分支)和B超群(红色分支),其中A超群包含埃及伊蚊中2条序列和白纹伊蚊中10条序列,与wAlbA亚群聚为一支,与NCBI中美国白纹伊蚊的AF020058.1菌株序列、我国广东省致倦库蚊(Culex quiquefasciatus=Cx. pipiens quiquefasciatus)的KJ140133.1和江苏省白纹伊蚊的MW718057.1菌株以及西班牙白纹伊蚊的MT569514.1菌株序列相似度高;B超群包含埃及伊蚊中的15条序列和白纹伊蚊中的79条序列,与wAlbB亚群聚为一支,在NCBI上共有3个序列相似度高的菌株,分别为斯里兰卡白纹伊蚊的KY523669.1菌株、广东省骚扰阿蚊(Armigeres subalbatus)的KJ140130.1菌株、美国白纹伊蚊的AF020059.1菌株。见图 1。
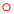
2.5 埃及伊蚊和白纹伊蚊沃尔巴克氏体wsp基因单倍型分析
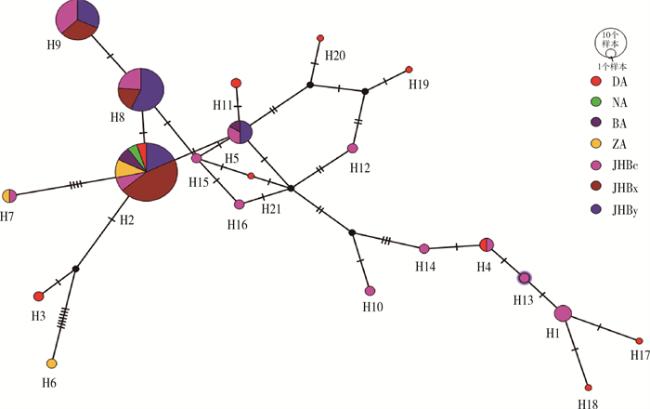
2个伊蚊种群自然感染沃尔巴克氏体所获106条沃尔巴克氏体wsp基因序列,使用Popart 1.7软件的TCS network构建单倍型网络图,见图 2。基因序列分析得到沃尔巴克氏体的多态性位点为26个,单倍型多样性(haplotype diversity,Hd)为0.794。共分为21个单倍型,包括5个埃及伊蚊和白纹伊蚊共享单倍型和16个独享单倍型(埃及伊蚊独享4个,白纹伊蚊独享12个),埃及伊蚊中共享单倍型和独享单倍型的样本比为13∶4,白纹伊蚊中为74∶15。H2为最大的一个共享单倍型,为39条wsp基因序列共享,H2中埃及伊蚊和白纹伊蚊的样本比为8∶31;其余共享单倍型中,H4中埃及伊蚊和白纹伊蚊样本比为1∶1,H5中为1∶5,H8中为2∶19,H9中为1∶18。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
表4 云南省景洪市埃及伊蚊和白纹伊蚊中沃尔巴克氏体wsp基因单倍型信息Tab. 4 Information on Wolbachia surface protein gene (wsp) haplotypes in Aedes aegypti and Ae. albopictus in Jinghong, Yunnan Province |
| 超群 | 单倍型 | 基因序列数 | 宿主 |
| A | H1 | 1 | 埃及伊蚊 |
| H4 | 2 | 埃及伊蚊、白纹伊蚊 | |
| H10 | 1 | 白纹伊蚊 | |
| H13 | 1 | 白纹伊蚊 | |
| H14 | 1 | 白纹伊蚊 | |
| H17 | 1 | 白纹伊蚊 | |
| H18 | 2 | 白纹伊蚊 | |
| B | H2 | 39 | 埃及伊蚊、白纹伊蚊 |
| H3 | 1 | 埃及伊蚊 | |
| H5 | 6 | 埃及伊蚊、白纹伊蚊 | |
| H6 | 1 | 埃及伊蚊 | |
| H7 | 1 | 埃及伊蚊 | |
| H8 | 21 | 埃及伊蚊、白纹伊蚊 | |
| H9 | 19 | 埃及伊蚊、白纹伊蚊 | |
| H11 | 3 | 白纹伊蚊 | |
| H12 | 1 | 白纹伊蚊 | |
| H15 | 1 | 白纹伊蚊 | |
| H16 | 1 | 白纹伊蚊 | |
| H19 | 1 | 白纹伊蚊 | |
| H20 | 1 | 白纹伊蚊 | |
| H21 | 1 | 白纹伊蚊 |
3 讨论
本次调查发现景洪市埃及伊蚊沃尔巴克氏体自然感染率为3.54%,与Zhang等[11]2018年在云南省西双版纳州景洪市捕获的埃及伊蚊中沃尔巴克氏体感染率为6.80%(20/292)相比较低,可能与景洪市登革热暴发频繁,全城大范围使用卫生杀虫剂灭蚊,埃及伊蚊种群更替有关,与Chao和Shih[12]2023年调查的台湾地区埃及伊蚊沃尔巴克氏体平均感染率3.33%基本一致。此外,其他研究显示印度东北部埃及伊蚊中沃尔巴克氏体感染率为38%[13],澳大利亚凯恩斯市为37.26%[14],菲律宾为16.80%[15],而斯里兰卡[16]、瓜德罗普岛[17]、美国加利福尼亚州[18]、巴西马拉尼昂州[19]等地区野生埃及伊蚊未发现感染沃尔巴克氏体,上述研究提示埃及伊蚊沃尔巴克氏体自然感染率各地区不同,总体偏低。景洪市白纹伊蚊沃尔巴克氏体自然感染率为92.71%,与我国福建省(90.5%)[20]、贵州省(92.9%)[21]、河南省(97.9%)[22],国外印度(85%)[13]、巴西(72%)[19]、西班牙(94%)[23]等报道结果基本一致,提示自然界中白纹伊蚊沃尔巴克氏体感染率均较高。本次调查显示景洪市埃及伊蚊沃尔巴克氏体自然感染率远低于白纹伊蚊,原因可能是沃尔巴克氏体广泛分布于自然界昆虫和其他节肢动物的细胞内,宿主间频繁物理接触均为沃尔巴克氏体的水平转移提供了条件,而埃及伊蚊属于家栖型蚊种,白纹伊蚊属于半家栖型蚊种[24],白纹伊蚊较埃及伊蚊有更多的机会与沃尔巴克氏体形成共生。
本研究中同一世代的埃及伊蚊成蚊与幼蚊、雌蚊与雄蚊沃尔巴克氏体自然感染率无统计学差异,白纹伊蚊的研究结果也与其一致,提示虫态改变、性别对沃尔巴克氏体自然感染率无影响,与吕锡宏等[25]2024年研究上海市松江区白纹伊蚊成蚊与幼蚊、不同性别白纹伊蚊自然感染率存在差异的结果不一致,其研究显示白纹伊蚊雌蚊和雄蚊的沃尔巴克氏体检出率分别为50.7%(76/150)和3.7%(1/27),两者存在差异;白纹伊蚊成蚊和幼蚊的沃尔巴克氏体总检出率分别为43.5%(77/177)和70.4%(190/270),两者存在差异。原因可能是吕锡宏等的研究样本伊蚊成蚊和幼蚊分别采自不同孳生环境,其差异可能混杂有不同孳生环境的影响,而本研究为同一世代的幼蚊和由其羽化的成蚊,蚊虫的世代差异是否能导致沃尔巴克氏体自然感染率的差别,需进一步探究。
本研究发现埃及伊蚊和白纹伊蚊中存在wAlbA、wAlbB以及wAlbA+wAlbB共感染3种情况。Zhang等[11]发现2018年景洪市埃及伊蚊、王政艳等[21]发现2020年贵州省白纹伊蚊、吕锡宏等[25]发现2024年上海市白纹伊蚊均存在wAlbA+wAlbB共感染的情况,提示同种蚊种感染不同型沃尔巴克氏体普遍存在。景洪市白纹伊蚊中wAlbA+wAlbB共感染构成比高于埃及伊蚊,可能也是因为白纹伊蚊有更多的机会与沃尔巴克氏体形成共生。基于wsp基因通用引物检测沃尔巴克氏体自然感染,扩增产物通过比对可进一步确定亚群,但无法分析共感染情况,而基于wAlbA和wAlbB分型检测时,可分析wAlbA+wAlbB共感染情况,但无法检测其他亚群,研究中应根据实际情况选择检测对象。
本研究中埃及伊蚊17条wsp基因进化分析显示其基因型是wAlbA和wAlbB,2条属于wAlbA,15条属于wAlbB,未发现其他亚群。同样,白纹伊蚊89条wsp基因序列,仅发现wAlbA和wAlbB,以wAlbB为主,未发现其他亚群。与杨菂子等[26]2019年调查广州市白纹伊蚊、致倦库蚊和骚扰阿蚊普遍感染沃尔巴克氏体,检出wAlbA和wAlbB结果一致,但与燕清丽等[27]2018年调查发现江苏省淮安市蚊种自然感染沃尔巴克氏体,存在Mors、Dei和Pip基因型的结果不同,可能是因为本次调查以埃及伊蚊和白纹伊蚊蚊种为主,而淮安市调查的蚊种以三带喙库蚊、骚扰阿蚊和淡色库蚊(Cx. pipiens pallens)为主,蚊种不同导致自然感染沃尔巴克氏体的亚群不同,但需进一步开展研究证实。
本研究中106条wsp序列可分为21个单倍型,单倍型多样性为0.79,与刘倩倩等[28]和燕清丽等[27]研究中江苏省蚊虫自然感染沃尔巴克氏体的单倍型多样性为0.924相比偏低,可能是由于研究蚊种不同,导致蚊虫自然感染沃尔巴克氏体基因的单倍型多样性不一致。21个单倍型中,H2、H4、H5、H8和H9单倍型为埃及伊蚊和白纹伊蚊共有,提示蚊种间可能存在基因交流。H10~H21为白纹伊蚊的12个特定单倍型,其中H10~H19为雌蚊的10个特定单倍型,H20为雄蚊的特定单倍型,H21为幼蚊的特定单倍型,白纹伊蚊雌蚊单倍型数远多于雄蚊和幼蚊,提示白纹伊蚊雌蚊基因可能存在更多的分化,从而容易衍生出较多的单倍型。H1、H3、H6、H7为埃及伊蚊的特定单倍型,但由于本次从埃及伊蚊所获wsp基因序列较少,不同虫态之间单倍型分化未呈现明显规律。从单倍型看,埃及伊蚊沃尔巴克氏体单倍型远少于白纹伊蚊,是否与白纹伊蚊感染率高于埃及伊蚊有关,需进一步探讨。
综上所述,景洪市埃及伊蚊和白纹伊蚊自然感染沃尔巴克氏体的基因分型为wAlbA和wAlbB,存在wAlbA+wAlbB共同感染。埃及伊蚊沃尔巴克氏体自然感染率远低于白纹伊蚊,虫态和性别未对伊蚊的沃尔巴克氏体自然感染产生影响。系统进化分析沃尔巴克氏体基因存在一定的进化和变异,白纹伊蚊沃尔巴克氏体单倍型较埃及伊蚊丰富。本次研究发现景洪市埃及伊蚊沃尔巴克氏体感染率较低,未来使用基于沃尔巴克氏体的种群压制和种群替换策略防控埃及伊蚊成为可能。